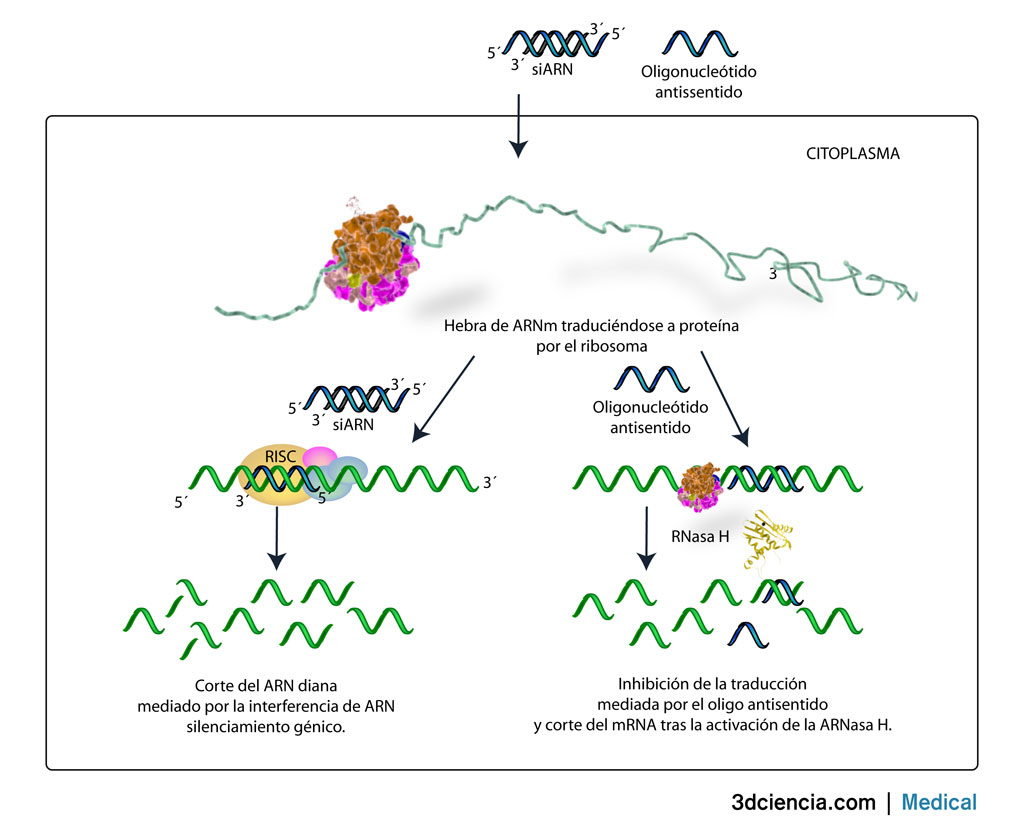
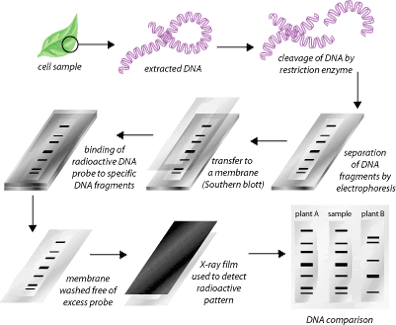
 The Southern blot es un método para
sondear la presencia de una secuencia específica de ADN en una muestra de ADN.
Muestras de ADN antes o después de la digestión de la enzima de restricción se
separan por electroforesis en gel y luego transferidas a una membrana por Blot
a través de la acción capilar. La membrana entonces está expuesta a una sonda
de ADN etiquetada que tiene una secuencia base de complemento a la secuencia de
ADN de interés. Protocolos más originales utilizan etiquetas radiactivas, ahora
existen alternativas sin embargo no radiactivas. Southern Blot es menos
utilizada en Ciencias de laboratorio debido a la capacidad de otras técnicas,
tales como PCR, para detectar secuencias específicas de ADN de muestras de ADN.
Estas manchas se siguen utilizando para algunas aplicaciones, sin embargo, como
el número de copia de transgen en ratones transgénicos, o en la ingeniería de
líneas de células madre embrionarias de gene knockout de medición.
The Southern blot es un método para
sondear la presencia de una secuencia específica de ADN en una muestra de ADN.
Muestras de ADN antes o después de la digestión de la enzima de restricción se
separan por electroforesis en gel y luego transferidas a una membrana por Blot
a través de la acción capilar. La membrana entonces está expuesta a una sonda
de ADN etiquetada que tiene una secuencia base de complemento a la secuencia de
ADN de interés. Protocolos más originales utilizan etiquetas radiactivas, ahora
existen alternativas sin embargo no radiactivas. Southern Blot es menos
utilizada en Ciencias de laboratorio debido a la capacidad de otras técnicas,
tales como PCR, para detectar secuencias específicas de ADN de muestras de ADN.
Estas manchas se siguen utilizando para algunas aplicaciones, sin embargo, como
el número de copia de transgen en ratones transgénicos, o en la ingeniería de
líneas de células madre embrionarias de gene knockout de medición.
Western Blot
Anticuerpos contra la mayoría de las
proteínas se puede crear inyectando pequeñas cantidades de la proteína en un
animal como un ratón, conejos, ovejas o burro (anticuerpos policlonales) o
producido en cultivos celulares (anticuerpos monoclonales). Estos anticuerpos
pueden utilizarse para una variedad de técnicas analíticas y preparativas.
 En western Blot, proteínas en primer
lugar se separan por tamaño en un gel fino entre dos placas de vidrio en una
técnica conocida como SDS-PAGE (electroforesis en gel de poliacrilamida sodio
Dodecil sulfato). Las proteínas en el gel luego son transferidas a un PVDF,
nitrocelulosa, nylon o otra membrana de soporte. Esta membrana, a continuación,
puede ser sondeada con soluciones de anticuerpos. Los anticuerpos que se unen
específicamente a la proteína de interés, a continuación, pueden visualizarse
por una variedad de técnicas, incluyendo productos coloreados,
quimioluminiscencia o autorradiografía.
En western Blot, proteínas en primer
lugar se separan por tamaño en un gel fino entre dos placas de vidrio en una
técnica conocida como SDS-PAGE (electroforesis en gel de poliacrilamida sodio
Dodecil sulfato). Las proteínas en el gel luego son transferidas a un PVDF,
nitrocelulosa, nylon o otra membrana de soporte. Esta membrana, a continuación,
puede ser sondeada con soluciones de anticuerpos. Los anticuerpos que se unen
específicamente a la proteína de interés, a continuación, pueden visualizarse
por una variedad de técnicas, incluyendo productos coloreados,
quimioluminiscencia o autorradiografía.
Northern Blot

The northern blot se utiliza para
estudiar los patrones de expresión de un tipo específico de la molécula de ARN
como comparación relativa entre un conjunto de diferentes muestras de ARN. Es
esencialmente una combinación de desnaturalización electroforesis en gel RNA y
una mancha. En este proceso de ARN está separado basado en tamaño y es
transferido a una membrana que luego es sondeada con un etiquetado como
complemento de una secuencia de interés. Los resultados pueden visualizarse a
través de una variedad de formas dependiendo de la etiqueta utilizada; Sin embargo,
la mayoría como resultado la revelación de bandas que representa el tamaño del
ARN detectado en la muestra. La intensidad de estas bandas está relacionada con
la cantidad del objetivo del RNA en las muestras analizadas. El procedimiento
es utilizado comúnmente para estudiar cuándo y cuánto expresión génica ocurre
midiendo cuánto de ese ARN está presente en diferentes muestras. Es una de las
herramientas más básicas para determinar en qué momento y bajo qué condiciones,
algunos genes se expresan en tejidos vivos.